Research
In the Green Lab we study sub-cellular to supra-cellular biology, from molecule to man. We are elucidating the machinery that mechanically and chemically couples cells in tissues, and determining how interference with this machinery can lead to human disease. Alteration in cell-cell coupling not only changes tissue architecture, but also alters paracrine communication between different cell types, including cells of the immune system.
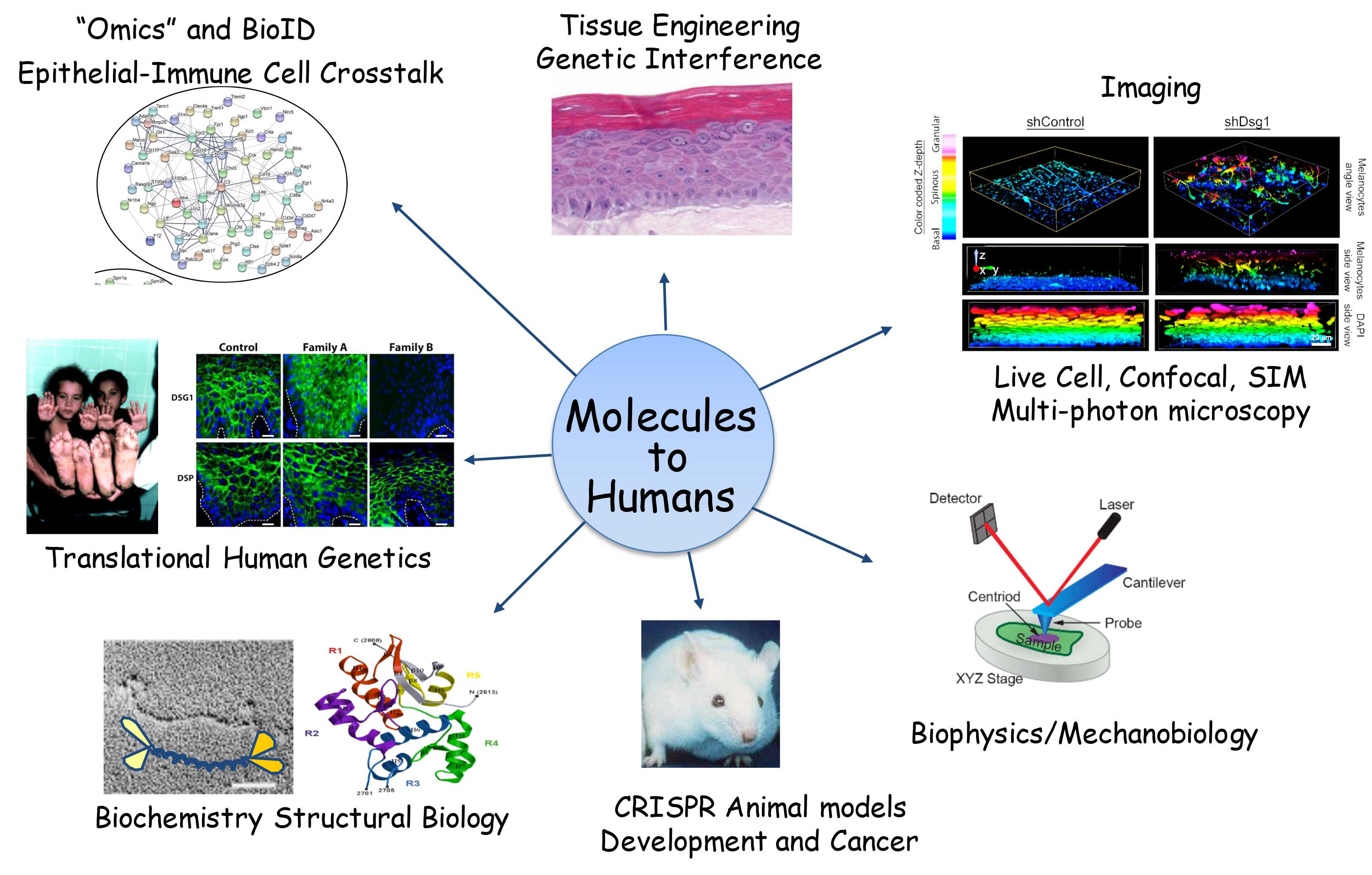
This figure illustrates different approaches used in the lab, starting at the left and moving clockwise with:
- studies of human desmosome-related disease
- transcriptomic and proteomic approaches to identify new interaction partners and functional changes in mRNA/protein expression
- regeneration of human epidermis in vitro from isolated cells
- live and fixed imaging of cells and tissues
- biophysical approaches to study mechanical signaling
- generation of animal models of human genetic disease and cancer including melanoma
- biochemical and structural biology approaches
Through these approaches, we learn how human mutations alter protein dynamics and function within cells, and how mutations change cell communication within tissues to alter tissue function and immune responses. (images from: Norgett, et al. 2000. Hum. Mol. Gen.; Samuelov, et al. 2013. Nat. Genet.; Choi, et al. 2002. Nat. Struct. Biol.; O’Keefe, et al. 1989. J. Biol. Chem.)
Choose a topic below to learn more about our work: